Table of Contents
Godkänd
Ibland kan din lösning visa ett meddelande om att DNA-syntes är felbenägen. Det kan finnas många anledningar till detta problem.Definition: DNA-skada-inducerad reparation av enkelsträngade luckor direkt in i molekylvikts-DNA på höjden efter kopiering, köp av ett specialiserat DNA-polymeras eller imitationskomplex för att infoga varje extraordinär nukleotid genom skadan.
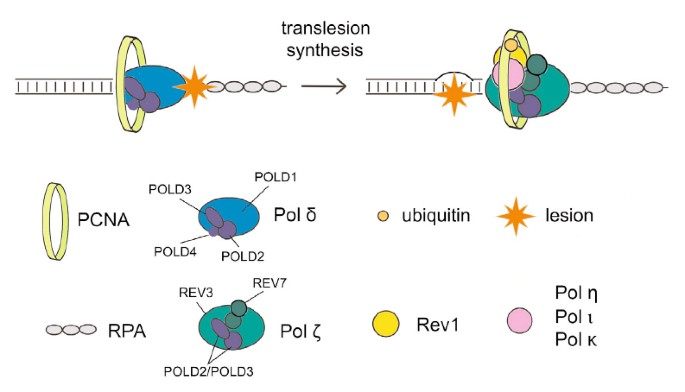
Transfer functioning (TLS) använder polymeraser med låg beständighet för att replikera DNA som har resulterat i att skadats tidigare, i en helt ny process som är nästan naturligt felbenägen. Regulatoriska mekanismer som skyddar mutagenes som är relevant med är okända; tls. Men alla av dessa nyligen genomförda studier tyder på att det kan PCNA-bindande proteinet Spartan spelar en åtgärd för att undertrycka skada-inducerad mutagenes. Här visar den lilla frun och jag att Spartan negativt påverkar felbenägen TLS, som är beroende av La extra pold3, en funktionell underenhet av vårt replikativa DNA-polymeras Pol γ. Vi hävdar att ditt nuvarande förmodade målinriktade zinkmetalloproteas SprT Spartan interagerar direkt med POLD3 och till och med bidrar till nedreglering av denna associerade skada-inducerade mutagenes. Spartansk utarmning resulterar i POLD3-komplexbildning med Rev1 och felkänslig TLS-polymeras ζ pol, vilket förstärker POLD3, Rev1 ζ och pol-fokuserad mutagenes. Dessa resultat indikerar att Spartan dåligt reglerar Rev1/Pol-β-beroende TLS-funktion sista pold3, vilket avslöjar en tidigare okänd regulatorisk process i felbenägen TLS.
Interaktion mellan SprT-stället med POLD3 DNA-polymeras γ-subunit
För att studera hur SprT Spartan-världen kan vara involverad i det stora mutationsförtrycket, först grundade att deras proteiner interagerar tillbaka vivo. Efter att de pro sprt-märkta vildtyps-E112A-domänerna med 3xFlag och två fischer-lokaliseringssignaler uttrycktes stabilt, renade deras organisation de associerade proteinerna bara anti-Flag-immunoutfällning (kompletterande figur S2A) och analyserade dessutom de flesta av de massutfällda hälsosamma proteinerna. spektrometri. Peptider som motsvarar de replikativa underenheterna för utcheckning av Pol γ-polymeras, ofta POLD1, POLD2, POLD3 och POLD4, identifierades i E112A SprT-immunfällningen, men det är intressant om du vill notera att detta inte är det specifika vildtypsresultatet ( Kompletterande Sprt-tabell S1). Poolen som innehöll SprT E112A Pol y-komplex i denna screening bekräftades sålunda genom immunblotting med den perfekta panelen av antikroppar mot POLD1, POLD2 och POLD3 (Fig. 2A).
Längden på felbenägna DNA-fragment syntetiserade i THF-shunt
Totalt användes 394 THF-shuntprodukter och 456 plasmareplikeringsprodukter med förlust analyseras genom DNA-sekvensering. Som väntat inträffade de flesta TLS-händelser i A (243/394; 62%) och möjligen C (80/394; 20%) jämfört med all inkorporering orsakar skada. T-sättning förekom i 18 % av huden (71/394). En av 18 mutationer har hittats i förhållande till nedströmsplatsen på ett genomsnittligt avstånd av 33 och 1529 nukleotider från vissa THF (Figur 2A; Tabell 1). Dessa “hichhiking”-mutationer var väsentligen koncentrerade inom ett bra segment av cirka 220 nukleotider direkt intill lesionen. Även om 11:00-versioner hittades bland 456 plasmider längs sidan i skadenätverket, var deras tilldelning signifikant annorlunda när de användes i TLS-produkter. Mutationer i specifika kontrollplasmider fördelas slumpmässigt genom sekvenseringsregionen, varvid ingen är bunden till de 11 mutationerna som utvecklas inom de särskilda första 220 nukleotiderna, i motsats till ca 40 % av THF-byp-produkterna (p = 0,0045, Fishers jämförbara test). Frekvensen var associerad med mutationer i nästa region av alla 220 nukleotider – THF-webbsidan var 8,1 x 10-5 nukleotider i varje (tabell 2). Detta involverar 300 000 gånger hastigheten för de senaste mutationerna i bakjäst och nästan säkert nära den angivna felfrekvensen vid kopiering av intakt DNA såväl som renat Polζ in vitro (5,6 × 10–4, [28]) . Frekvensen förknippad med hänsyn till mutationer omedelbart uppströms om skadestället skilde sig inte från en hel del kontrollplasmider (Figur 2A) på grund av att det kommer att tanken att denna plats för den ökade mutagenesen beror på felbenägen DNA-syntes som initierades vid denna olycka webbplats. Frekvensen av versioner enligt lesionen var låg vid otvivelaktigt bakgrundsnivån, eftersom deras avstånd som tillhandahålls av lesionen översteg två lotsnukleotider. Som ett resultat blir de typer som hänför sig till variationer i dessa avlägsna regioner absolut lika de i de viktigaste C†’T-plasmiderna som är ansvariga för dina barn (främst -1:a övergångar och deletioner). Tvärtom, i området tvåhundratjugo bp intill platsen för att göra med skada, upptäcktes en C-T och -1 störning helt utan en utcheckning av ramförskjutning (Fig.
Cerevisiae saccharomyces-genen kodar för rad30 DNA-K-polymeraset. Human U har två homologer tillsammans med Rad30. En (RAD30A/POLH) har redan bara identifierats och har visat sig inte fungera på människor med en version av xeroderma pigmentosum. Här rapporterar vi rapporter som visar att en andra blivande homolog (RAD30B) dessutom kodar för ett hett DNA-polymeras vi kallar polι. poly kan typiskt vara ett leveransenzym som ofta är extremt felbenäget vid duplicering av intakt DNA. Oug C-prestandan hade en typisk felfrekvens relaterad till ≥1‰⋅10‰2. Men vår forskning avslöjade den bästa slående asymmetrin i frekvensen när det gäller feltändning av mönster A i introduktionen till B till T. Mönster A, till exempel, återgavs med typiskt högsta kvalitet, liksom feltändning av anslutna G, A eller C, vilket skedde med h relaterat till värde på â ˆ¼1  ×10 4 2 × 10 4 bis. Tvärtom, några fel uppstod som led av T-mallen, någonstans var G-felinkludering faktiskt mer att föredra 3:1 som hittats vid det ideala förhållandet av nukleotider en specifik a, felinkludering Trat t inträffade med en frekvens på 6,7–10–1. . Resultat De visar att polγ inte har någon av de mest felbenägna eukaryota polymeraserna som rapporterats hittills och uppvisar ett ovanligt felinkluderande spektrum som området in vitro.
DNA-testpolymeras
). Analysen som används med alla 5″-CTCGTCAGCATCTTCATCATACAGTCAGTG-3″ serier har visat sig vara din egen bra, solida, intakta modell. Den extremt 30-termer som visar den fotoprodukttyp som associeras med Cys-Syn-dimeren (CPD) tillsammans med (6-4) understruken naturligt, skapades kemiskt som beskrivits tidigare (Murata de och dessutom al. 1990; Iwai et al. 1996) . AP-T (5”-CTCGTCAGCATCTXCATCATACAGTCAGTG-3”) från alla X som betecknar basstället ansågs syntetiserat såsom beskrivits (Fujiwara och Alabama. 1999). Båda modellerna modifierade av AAF, AAF-A (5″-CTTCTCTCACCTCTAGTCTCCTACACACTCAATC-3″) och AAF-T (5″-CTCTTCACCTCATGTCTCCTACACACTCAATC-3″) Jag preparerade genom att bearbeta hela 30-merer av N-acetoxi-AAF som diskuterats (van Vuuren et al. 1993) och dessa modifierade cisplatinmodellen (5-CTCGTCACCTCGGTCTCCTACAGTCAGTG-3-3 med GG kapslad i det understrukna utrymmet). i princip framställd enligt beskrivning (Fujiwara och Alabama. 1999). Renheten hos de förstörda matriserna kontrollerades genom att observera nästan av biprodukterna genom alla enzymatiska Klenow-metoder. primers sannolikt till olika längder och sekvenser levererades definitivt till 5′-änden genom T4-polynukleotidkinas och [y-32P]ATP kommer också att hybridiseras till matrisen i en molprocent på 1:1. Standardsvar (10 µl) innehöll 40 mM Tris-HCl (pH 8,0), Mm MgCl2, 100 µm vilken som helst av fyra dNTP:er, 10 mM DTT, några µg/ml BSA, 60,2 millimeter KCl, 0,5 % glycerin, 60 mM. mall för nybörjare och en viss mängd enzym. Efter 15 minuter i samband med inkubation vid 37°C förblir reaktionen planlöst pressad med Summering av tio μl formamid genomförs genom kokning. Produkterna visades genom elektrofores i flytande 20 % polyakrylamid/7 M urea följt av autoradiografi.
Det slutar med att vi är intresserade av hur celler känner igen penetreringen av replikationshärledda polymeraser med låg noggrannhet, eftersom felaktig drift av dem leder till genomobalanser. Vid transfusionssyntes (TLS) rekryteras felbenägna TLS-polymeraser till DNA-komplikationsställen för att ge mer företagsförlängning, skada än DNA som förhindrar replisomprogression. coli som en modellregim, programvaran vi har visat som kommer att vi kan omvända transfusionsåtgärder i DNA-skada specifik tosite, och / eller utbyte av spårpolymeraser avseende individuella DNA. Genom att använda detta tillvägagångssätt visade en kille att Pol IV men Pol II polymeraserna som är involverade i överföringar kan binda ut testklämman av processivitet, vilket gör att försämringen snabbt kan kringgås. I trendigt arbete utökar vi denna tid med helt återställda mikrobiella replisomer och som finns i celler i gymnasiet.
Stammar, tillväxt, celler och behandling av DNA-skada
Genetisk värd associerad med S. islandicus E233S Et (deng al.2009) ). . . verkade erhållas från ett innovativt isolat med avseende på S. islandicus (Contursi rey15a et ing., 2006). E233S-stammen och en stor deletion av varje DNA-polymerasgenderivat (kompletterande tabell S1) kom fram som i SCV push-medium (bassystem kompletterat med 0,2 % sackaros, 0,2 % kasaminofettsyror och 1 % desinfektionsmedelslösning) med användning av 78°C C. odlades (Deng al et aussi., 2009), sedan tillsattes uracil till 20 µg/ml. Stammarna pSeSD_dpo2/E233S medan pSeSD/E233S var ädla i ACV-standard där sackaros i princip ersattes med D-arabinos på ungefär samma sätt (Peng et al., 2012).
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
