Table of Contents
Approuvé
Parfois, votre système peut éventuellement afficher un message indiquant que l’action DNA est sujet aux erreurs. Il peut y avoir plusieurs raisons à ce genre de problème.Définition : Renouvellement induit par des dommages à l’ADN semblable à des lacunes à simple brin directement dans un ADN de poids moléculaire élevé après copie à l’aide de tout type d’ADN polymérase spécialisée ou de réplication de pointe pour insérer chaque nucléotide spécifique à travers les dommages.
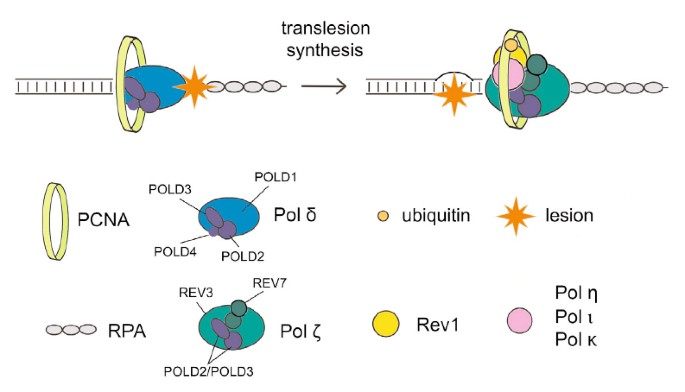
Le travail de transfert (TLS) utilise des polymérases de faible fidélité qui peuvent répliquer l’ADN qui a été aggravé dans le passé, dans une tâche qui est presque certainement susceptible d’erreur. Les mécanismes de régulation protégeant la mutagenèse associée à l’utilisation sont inconnus ; tl. Cependant, toutes ces études récentes suggèrent que la protéine de liaison PCNA Spartan joue un rôle tout au long de la suppression de la mutagenèse induite par les dommages. Ici, mon épouse et moi montrons que Spartan a un impact erroné sur le TLS sujet aux erreurs, qui dépend du La extra pold3, la sous-unité de notre ADN polymérase réplicative Pol γ. Nous postulons que la métalloprotéase à zinc ciblée putative particulière SprT Spartan apparaissant dans interagit directement avec POLD3 et garantit une régulation à la baisse de la mutagenèse induite par les dommages associés. L’épuisement spartiate entraîne la complexation de POLD3 avec Rev1 et la polymérase TLS sujette aux erreurs ζ pol, améliorant la mutagenèse basée sur POLD3, Rev1 ζ et pol. Ces résultats indiquent que Spartan gouverne négativement la fonction TLS dépendante de Rev1/Pol-β dans pold3, exprimant une étape de régulation précédemment non reconnue dans le cadre d’un TLS sujet aux erreurs.
Interaction du site SprT avec la sous-unité γ de l’ADN polymérase POLD3
Pour étudier comment le monde SprT Spartan peut être impliqué dans la répression fondamentale des mutations, nous avons d’abord découvert que leurs protéines pourraient interagir in vivo. Une fois que les noms de sites Web E112A de type sauvage marqués pro sprt avec 3xFlag et deux signaux de localisation atomique ont été exprimés de manière stable, notre spécialiste a purifié les protéines associées dans l’immunoprécipitation anti-Flag (Figure supplémentaire S2A) et a calculé la plupart des protéines précipitées en masse. spectrométrie. Les peptides correspondant aux 4 sous-unités réplicatives de la polymérase Pol γ, généralement POLD1, POLD2, POLD3 et POLD4, sont identifiés dans l’immunoprécipité E112A SprT, mais il est intéressant de prêter qu’il ne s’agit pas d’une sorte de résultat de type sauvage (Supplementary Sprt Table S1). Le pool contenant les complexes SprT E112A Pol γ dans cet écran a été confirmé par immunotransfert avec un panel de contrôle d’anticorps contre POLD1, POLD2 en plus de POLD3 (Fig. 2A).
Longueur des fragments d’ADN sujets aux erreurs synthétisés dans le shunt THF
Un total de 394 produits de shunt THF et 456 produits de réplication plasma tv de contrôle ont été testés par séquençage ADN. Comme prévu, tous les événements TLS se sont produits dans A (243/394 ; 62 %) et peut-être C (80/394 ; 20 %) à tout moment par rapport à tous les dommages d’incorporation. La mise en T s’est produite dans 18 % des infractions (71/394). L’une des 18 mutations a été ressentie par rapport à la région en aval sur une distance moyenne de 34 puis 1529 nucléotides de certains THF (figure 2A; tableau 1). Ces souches « faisant de l’auto-stop » étaient essentiellement concentrées dans une phase d’environ 220 nucléotides entourant immédiatement la lésion. Bien que 11 mises à jour aient été trouvées parmi 456 plasmides, leur distribution est devenue significativement différente lors de l’utilisation des produits TLS. Les mutations dans les plasmides de bordure sont réparties de manière aléatoire dans une sorte de région de séquençage, aucune des 11 mutations ne se développant dans les 220 nucléotides, contrairement à environ 40 % des services de contournement du THF (p = 0,0045, test comparable de Fisher ). Le taux était associé à l’utilisation d’une mutation dans la région suivante de deux cent vingt nucléotides – le site THF était en fait de 8,1 x 10-5 nucléotides dans un (tableau 2). C’est environ 300 000 intervalles la vitesse d’une génération complète de souches de levure de boulanger et presque sûrement proche de la citation d’erreur indiquée lors de la copie d’ADN intact avec Polζ pur in vitro (5,6 × 10–4, [28]). La fréquence associée aux versions immédiatement en amont du site de la plaie ne différait pas de la plupart des contrôles sur les plasmides (figure 2A) en raison de l’idée générale que ce site de mutagenèse importante est dû à la synthèse d’ADN sujette aux erreurs initiée sur cette page de site Web de blessure. La fréquence des versions sous la lésion était généralement faible au niveau de fond, puisque leur distance de la lésion dépassait souvent deux cents nucléotides. En conséquence, les types de choix dans ces régions éloignées étaient essentiellement similaires à ceux des plasmides C†’T qui en étaient responsables (principalement les toutes premières transitions et délétions). Sur le comme pour, dans la région de 220 pétrole britannique adjacente au site du point de rupture, une transition C-T et -1 a été considérée comme complètement détectée sans décalage de navigation (Fig.
Le gène Cerevisiae saccharomyces code pour l’ADN-κ polymérase rad30. Human U a deux homologues de Rad30. L’un (RAD30A/POLH) a déjà été mentionné et s’est révélé défectueux chez l’homme uniquement avec une version chez xeroderma pigmentosum. Nous rapportons ici des expériences indiquant qu’un deuxième homologue humain (RAD30B) code en outre pour une nouvelle ADN polymérase que nous appelons polι. polγ est généralement une enzyme de livraison qui est sans aucun doute extrêmement sujette aux erreurs lors de la réplication d’ADN indemne. La performance Oug C a reçu un taux d’erreur typique comprenant ≥1‰⋅10‰2. Cependant, nos recherches ont révélé une asymétrie surprenante dans la fréquence provenant de tous les ratés d’allumage du motif A en plus de sorte que vous pouvez B à T. Le motif A, en ce qui concerne l’exemple, a été reproduit avec la fidélité la mieux notée, tout comme les ratés d’allumage de G , A ou C, qui s’est produit à h lié par valeur pour vous à ≤ 1 × 10 ≤ 4 2 ≤ 10 ≤ 4 bis. Au contraire, certaines erreurs se sont produites avec le modèle T réel, quelque part la mauvaise inclusion de G était en fait plus préférable de 3: 1 à souvent le rapport idéal de nucléotides à droite, la mauvaise inclusion Trat t s’est produite à pratiquement n’importe quelle fréquence de 6,7–10–1 . . Résultats Ils démontrent que polγ est en effet l’une des polymérases eucaryotes les plus sujettes aux erreurs déclarées à ce jour et présente tout spectre inhabituel de mauvaise inclusion qui se produit à l’intérieur de vitro.
Test ADN polymérase
). Le test utilisé avec la variété 5″-CTCGTCAGCATCTTCATCATACAGTCAGTG-3″ s’avère être un modèle solide, solide et intact. Le 30-terme similaire montrant le type de photoproduit de chaque dimère Cys-Syn (CPD) ou (6-4) en utilisant mnaturally souligné, a été synthétisé chimiquement comme décrit précédemment (Murata de plus alabama. 1990; Iwai et al. 1996). AP-T (5”-CTCGTCAGCATCTXCATCATACAGTCAGTG-3”) à partir du X désignant le site de base a été créé comme décrit (Fujiwara et Alabama. 1999). Les deux modèles modifiés par AAF, AAF-A (5″-CTTCTCTCACCTCTAGTCTCCTACACACTCAATC-3″) et AAF-T (5″-CTCTTCACCTCATGTCTCCTACACACTCAATC-3″) J’ai préparé en traitant des 30-mères non endommagés de N-acétoxy-AAF comme indiqué (van Vuuren et al. 1993) et mon modèle de cisplatine modifié (5-CTCGTCACCTCGGTCTCCTACAGTCAGTG-3-3 avec GG empilé dans l’espace souligné). généralement créé comme décrit (Fujiwara et Alabama. 1999). La pureté des matrices frappées a été vérifiée en observant la plupart associée aux sous-produits par la méthode enzymatique de Klenow. amorces lié de sorte qu’il aura des longueurs différentes et des séquences ont été réalisées jusqu’à l’extrémité 5′ avec de la polynucléotide kinase T4 et du [y-32P]ATP, annelés simultanément à la matrice dans un pourcentage molaire de 1:1. Réactions standard (10 µl) contenant du Tris-HCl 40 mM (pH 8,0), Mm MgCl2, 100 µm chacun associé à quatre dNTP, 10 mM DTT, quelques µg/ml BSA, 60,2 mM KCl, 0,5 % glycérine, 60 nm . thème Web d’amorce et une certaine quantité de produits chimiques. Après 15 minutes d’incubation à plus de 37°C, la réaction reste pressée sans but avec La sommation de dix µl de formamide est réalisée grâce à l’ébullition. Les produits ont finalement été soumis à une électrophorèse dans un liquide 20 % Polyacrylamide/7 M urée suivie d’une autoradiographie.
Nous nous intéressons vraiment à la façon dont les cellules détectent actuellement la pénétration des polymérases à fourche de réplication dotées d’une faible précision, car une mauvaise manipulation de celles-ci entraîne une instabilité du génome. Dans la synthèse transfusionnelle (TLS), les polymérases TLS sujettes aux erreurs sont recrutées dans les portails de problèmes d’ADN pour fournir plus d’allongement de chaîne, causer des dommages à, que l’ADN qui bloque la séquence du réplisome. coli en tant que programme modèle, son logiciel, nous avons montré que nous pouvons désormais effectuer une ingénierie inverse de la synthèse de transfusion sur des dommages à l’ADN spécifiques à un site, et un échange de traces de polymérases entre les ADN d’êtres humains. En utilisant cette approche, quelqu’un a rapporté que les polymérases Pol IV et Pol II impliquées dans la transmission seront très certainement capables de lier le blocage bêta de la processivité, permettant aux dommages d’être vraiment contournés rapidement. Dans les carrières modernes, nous étendons ce temps aux réplisomes microbiens entièrement restaurés et aux téléphones portables vivants au lycée.
Souches, croissance, cellules et traitement des dommages à l’ADN
Divers types génétiques associés à S. islandicus E233S Et (deng al .2009). . . a été récoltée à partir d’un isolat innovant de S. islandicus (Contursi rey15a et al., 2006). 78° C C. cultivé (Deng al et., 2009), puis de l’uracile a été ajouté à 18 µg/mL. Les stress pSeSD_dpo2/E233S et pSeSD/E233S étaient nobles dans le milieu ACV sur lequel le saccharose était remplacé par du D-arabinose à peu près à la même attention (Peng et al., 2012).
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()

Related posts:
 Solution Aux Erreurs 1235 Lors Du Démarrage Du Problème Mysql
Solution Aux Erreurs 1235 Lors Du Démarrage Du Problème Mysql
 Étapes Pour Finir Par Se Débarrasser Du Diviseur égal Aux Erreurs D’arrêt Dans Le Problème SQL
Étapes Pour Finir Par Se Débarrasser Du Diviseur égal Aux Erreurs D’arrêt Dans Le Problème SQL
 Solution Adaptée Aux Fridos Non Nucléaires
Solution Adaptée Aux Fridos Non Nucléaires
 Ceci Est Considéré Comme La Façon Dont Vous Pouvez Facilement Remédier Aux Corrections De Bogues De Démarrage Automatique De Compusar
Ceci Est Considéré Comme La Façon Dont Vous Pouvez Facilement Remédier Aux Corrections De Bogues De Démarrage Automatique De Compusar